|
Description
Ambystoma gracile adults may be either metamorphosed terrestrial salamanders or neotenic water-dwellers.
Metamorphosed adults measure 14 - 22 cm in total length (Boundy and Balgooyen 1988). A glandular ridge forms a rounded top edge on the tail; the lower edge of the tail is sharp (Petranka 1998; Snyder 1963). Conspicuous oval parotoid glands sit immediately behind the
eyes. The tail ridge appears rough due to concentrations of granular glands (Brodie and Gibson 1969). Both the parotoid and tail ridge glandular areas are pitted with openings of poison glands. Broad head, relatively small eyes. No tubercles on the underside of the feet (Stebbins 1985).
They are dark brown, gray, or black above and a lighter brown below (Petranka 1998; Snyder 1963). Sometimes terrestrial individuals have small, irregular, whitish or yellowish spots and blotches on their dorsum (see Comments section below).
Neotenic adults are strictly aquatic and retain gills. They may grow as large as 13 cm in snout-vent length and 26 cm in total length (Boundy and Balgooyen 1988). The parotoid glands of untransformed adults are less prominent than those of transformed adults (Licht and Sever 1993).
They are brownish to olive green in color, mottled with yellow and black. Distinct yellow spots sometimes dot the sides and tail. Ventral color varies from cream to pale or dark gray (Petranka 1998).
Both terrestrial and neotenic males become darker than females during the breeding season. Unmetamorphosed adult males have hypertrophied hindlimbs and feet, are less spotted, and have an enlarged glandular ridge on the tail (Snyder 1956).
Distribution and Habitat
Country distribution from AmphibiaWeb's database: Canada, United States U.S. state distribution from AmphibiaWeb's database: Alaska, California, Oregon, Washington Canadian province distribution from AmphibiaWeb's database: British Columbia
Inhabits the northwest Pacific coast of North America. Found from southeastern Alaska on May Island south to the mouth of the Gualala River, Sonoma County, California. Occurs from sea level to timberline, but not east of the Cascade Divide. Range includes Vancouver Island in British Columbia and Cypress, Whidbey, Bainbridge, and Vashon Islands in Washington (Snyder 1963).
The spotted forms occur in the northern portion of A. gracile's range (Petranka 1998; Snyder 1963; see Comments section below).
Populations with neotenic adults are widespread, but the frequency of gilled adults increases with altitude such that adults at low and intermediate altitudes are almost all terrestrial, while adults at very high elevations are mostly neotenic (Eagleson 1976).
Terrestrial adults live in mesic habitats ranging from grasslands to mesophytic forests. Terrestrial adults are mostly fossorial, and are usually active on the surface only during fall rains and spring migrations to their aquatic breeding sites. However, they can sometimes be found under logs and other surface debris outside of the breeding season (Petranka 1998; Efford and Mathias 1969). Life History, Abundance, Activity, and Special Behaviors
Terrestrial adults may defend themselves vigorously. When threatened, they may close their eyes, assume a head-butting position, and elevate the tail. They can secrete a sticky white poison from glands on the head, back, and tail, using head-butts and and tail-lashes to smear the poison onto the attacker (Stebbins 1985; Licht and Sever 1993). A ticking sound may accompany this defense (Licht 1973). Larvae and neotenic adults flee attack from natural predators such as trout and beetle larvae (Nussbaum et al. 1983).
This species breeds in permanent or semi-permanent waters, ranging from small, shallow ponds to large, deep lakes, and sometimes including side pools of slow streams. Breeding takes place in the spring; the exact timing varies with latitude and elevation (Brown 1976; Licht 1969; Licht 1975). Mating pairs of A. gracile enact an elaborate courtship ritual that seems to vary between different regions. Fertilization is internal; the female picks up a spermatophore deposited by the male (Licht 1969; Knudsen 1960). Length of embryonic development is positively correlated with water temperature and varies from 2-9 weeks (Petranka 1998). Larva
No morphological differences have been found to distinguish neotenic larvae from those that will transform (Snyder 1963).
Larvae belong to the pond type, having long gills, long toes, and a long dorsal fin (Stebbins 1985). Hatchlings average about 8 mm in snout-vent length (Licht 1975).
They have concentrated dark pigment along the base of the dorsal fin. They often have sooty blotches on the dorsum and may have yellow flecks or spots along the sides (Stebbins 1985).
Older larvae vary in color from dark brown to olive green or light yellow (Stebbins 1985).
Most larvae in lowland populations transform when 12-14 months old, in the spring following hatching (Eagleson 1976; Licht 1975). Larvae at high elevations transform in their third year (Eagleson 1976).
Hatchlings feed on zooplankton; older larvae eat a variety of prey, including annelids, mollusks, insect larvae, copepods, ostracods, and flatworms (Licht 1975; Efford and Tsumura 1973).
Larvae and neotenic adults flee attack from natural predators such as trout and beetle larvae (Nussbaum et al. 1983).
Trends and Threats
The status of Ambystoma gracile populations is unknown (Blaustein et al. 1994). No declines are apparent in Canada (Weller and Green 1997). In the Cascade Mountains of Washington, Aubry and Hall (1991) found A. gracile to be far less abundant in young forests than in old-growth forests. However, Corn and Bury (1991) surveyed Western Oregon and found little correlation between salamander abundance and stand age, so it remains unclear whether A. gracile requires old growth forest to thrive. Clear-cutting, however, seems to render the habitat unsuitable; Corn and Bury found no salamanders on recent clear-cuts. Petranka (1998) suggests leaving a forest buffer of 200-250 m surrounding breeding sites used by terrestrial adults to preserve optimal environmental conditions for local populations.
At an elevation of 183 m in the Cascade Mountains of Oregon, Blaustein et al. (1995) found A. gracile embryos to hatch with less mortality when shielded from UV-B radiation than when allowed to develop exposed to direct sunlight. This experiment was conducted in pond water 5-10 cm deep, which they claim is a common oviposition site for A. gracile in the Oregon Cascades. However, females
often lay their eggs at greater depths and often in murky water, where they would be protected from UV-B, so the ecological relevance of Blaustein et al.'s experiment is controversial. Whether UV-B actually harms A. gracile populations is unknown (Petranka 1998).
Because A. gracile breeds in relatively permanent bodies of water, Canadian populations may be unaffected by the increase in aridity predicted to occur in Canada due to global warming. However, decreased survival due to drying of breeding habitat may
still occur (Ovaska 1997).
A. gracile populations may be negatively affected by introduced predatory fish, such as trout and char. These fish can reduce or eliminate montane populations of ambystomatid salamanders (Burger 1950; Blair 1951; Sprules 1974; Taylor 1983). Tyler et al. (1998) tested the effects of trout on larvae of A. gracile and A. macrodactylum in artificial ponds and found that trout reduced both
survivorship and average snout-vent length, and also narrowed the range of habitat use for larvae of both species. Trout also reduced average body mass of A. gracile larvae. Relation to Humans
The poison secreted by terrestrial Northwestern Salamanders may irritate the skin of some people (Stebbins 1985).
Comments
The spotted and unspotted varieties were originally designated as separate subspecies, with Ambystoma gracile decorticatum (British Columbia Salamander) possessing spots and Ambystoma gracile gracile (Brown Salamander) lacking spots. The spotted form occurs in the northern part of the range for A. gracile. A dividing line of 51 degrees north latitude was recognized between the two subspecies, with A. g. decorticatum occurring north of the line and A. g. gracile living to the south (Snyder 1963). Morphological characters such as the presence of 3 instead of 4 phalanges on the fourth toe, more prominent parotoid glands, and rows of 4 rather than 2 teeth on the prevomers have been used to differentiate between the unspotted A. g. gracile and the spotted A. g. decorticatum (Dunn 1944). However, Titus (1991) found that these distinctions do not always hold true and that genetic variation does not always correlate well with subspecies, and he suggested that A. gracile should not be split into subspecies.
The specific epithet gracile is derived from the Latin term gracilis which means "slender" or "delicate" (Snyder 1963).
The subspecific name decorticatum is taken from the Latin de ("from") and cortex ("bark" or "cork"), meaning "like bark" (Snyder 1963).
See another account at californiaherps.com.
References
Aubry, K. B., and Hall, P. A. (1991). ''Terrestrial amphibian communities in the southern Washington Cascade Range.'' Wildlife and Vegetation of Unmanaged Douglas-fir Forests, General Technical Report PNW-GTR-285. Ruggiero, L. F., Aubry, K. B., Carey, A. B., and Huff, M. H., technical coordinators, eds., USDA Forest Service, Northwest Research Station, Olympia, Washington., 326-338.
Blair, A. P. (1951). "Notes on the herpetology of the Elk mountains, Colorado." Copeia, 1951, 239-240.
Blaustein, A. R., Edmond, B., Kiesecker, J. M., Beatty, J. J., and Hokit, D. G. (1995). "Ambient ultraviolet radiation causes mortality in salamander eggs." Ecological Applications, 5(3), 740-743.
Blaustein, A. R., Wake, D. B., and Sousa, W. P. (1994). "Amphibian declines: Judging stability, persistence, and susceptibility of populations to local and global extinctions." Conservation Biology, 8(1), 60-71.
Boundy, J. and Balgooyen, T. G. (1988). "Record lengths of some amphibians and reptiles from the western United States." Herpetological Review, 19, 27.
Brodie, E. D., Jr. and Gibson, L. S. (1969). ''Defensive behavior and skin glands of the Northwestern Salamander.'' Herpetologica, 25, 187-194.
Brown, H. A. (1976). ''The time-temperature relation of embryonic development in the Northwestern Salamander, Ambystoma gracile.'' Canadian Journal of Zoology, 54, 552-558.
Burger, W. L. (1950). ''Novel aspects of the life history of two Ambystomas.'' Journal of the Tennessee Academy of Sciences, 25, 252-257.
Corn, P. S. and Bury, R. B. (1991). ''Terrestrial amphibian communities in the Oregon Coast Range.'' Wildlife and Vegetation of Unmanaged Douglas-fir Forests, General Technical Report PNW-GTR-285. K. Ruggiero, B. Aubry, A. B. Carey, and M. H. Huff, technical coordinators, eds., USDA Forest Service, Pacific Northwest Research Station, Olympia, Washington., 304-317.
Dunn, E. R. (1944). ''Notes on the salamanders of the Ambystoma gracile group.'' Copeia, 1944, 129-130.
Dunn, E. R. (1954). ''The status of Siredon gracilis Baird.'' Copeia, 1954, 135-136.
Dyrkacz, S. (1981). "Recent instances of albinism in North American amphibians and reptiles." Herpetological Circular of the Society for the Study of Amphibians and Reptiles, 11, 1-31.
Eagleson, G. W. (1976). ''A comparison of the life histories and growth patterns of populations of the salamander Ambystoma gracile (Baird) from permanent low-altitude and montane lakes.'' Canadian Journal of Zoology, 54, 2098-2111.
Efford, I. E. and Mathias, J. A. (1969). "A comparison of two salamander populations in Marion Lake, British Columbia." Copeia, 1969, 723-736.
Efford, I. E. and Tsumura, K. (1973). ''A comparison of the food of salamanders and fish in Marion Lake, British Columbia.'' Transactions of the American Fisheries Society, 1, 33-47.
Henry, W. V., and Twitty, V. C. (1940). "Contributions to the life histories of Dicamptodon tenebrosus and Ambystoma gracile." Copeia, 1940, 247-250.
Knudsen, J. W. (1960). ''The courtship and egg mass of Ambystoma gracile and Ambystoma macrodactylum.'' Copeia, 1960, 44-46.
Licht, L. E. (1969). "Observations on the courtship behavior of Ambystoma gracile." Herpetologica, 25, 49-52.
Licht, L. E. (1973). "Behavior and sound production by the northwestern salamander Ambystoma gracile." Canadian Journal of Zoology, 51, 1055-1056.
Licht, L. E. (1975). "Growth and food of larval Ambystoma gracile from a lowland population in southwestern British Columbia." Canadian Journal of Zoology, 70, 87-93.
Licht, L. E., and Sever, D. M. (1993). "Structure and development of the paratoid gland in metamorphosed and neotenic Ambystoma gracile." Copeia, 1993, 116-123.
Nebeker, A. V., Schuytema, G. S, Griffis, W. L., and Cataldo, A. (1998). ''Impact of guthion on survival and growth of the frog Pseudacris regilla and the salamanders Ambystoma gracile and Ambystoma macrodactylum.'' Archives of Environmental Contamination and Toxicology, 35(1), 48-51.
Nebeker, A. V., Schuytema, G. S., and Ott, S. L. (1995). ''Effects of cadmium on growth and bioaccumulation in the Northwestern Salamander Ambystoma gracile.'' Archives of Environmental Contamination and Toxicology, 29(4), 492-499.
Nebeker, A. V., Shuytema, G. S., and Ott, S. L. (1994). "Effects of cadmium on limb regeneration in the Northwestern salamander." Archives of Environmental Contamination and Toxicology, 27(3), 318-322.
Nussbaum, R. A., Brodie, E. D., Jr., and Storm, R. M. (1983). Amphibians and Reptiles of the Pacific Northwest. University of Idaho Press, Moscow, Idaho.
Ovaska, K. (1997). ''Vulnerability of amphibians in Canada to global warming and increased solar ultraviolet radiation.'' Amphibians in Decline: Canadian Studies of a Global Problem. Green, D. M., eds., Herpetological Conservation, 206-225.
Petranka, J. W. (1998). Salamanders of the United States and Canada. Smithsonian Institution Press, Washington D.C. and London.
Richardson, J. P. M. and Adamson, M. L. (1988). ''Megalobatrachonema (Chabaudgolvania) waldeni n. sp. (Nematoda: Kathlaniidae) from the intestine of the Northwestern Salamander, Ambystoma gracile (Baird).'' Canadian Journal of Zoology, 66, 1505.
Slater, J. R. (1936). ''Notes on Ambystoma gracile Baird and Ambystoma macrodactylum Baird.'' Copeia, 1936, 234-236.
Snyder, R. C. (1956). ''Comparative features of the life histories of Ambystoma gracile (Baird) from populations at low and high altitudes.'' Copeia, 1956, 187-194.
Snyder, R. C. (1963). ''Ambystoma gracile.'' Catalogue of American Amphibians and Reptiles. American Society of Ichthyologists and Herpetologists, 6.1-6.2.
Sprules, W. G. (1974). ''The adaptive significance of paedogenesis in North American species of Ambystoma (Amphibia: Caudata): an hypothesis.'' Canadian Journal of Zoology, 52, 393-400.
Stebbins, R. C. (1985). A Field Guide to Western Reptiles and Amphibians. Houghton Mifflin, Boston.
Taylor, J. (1983). ''Orientation and flight behavior of a neotenic salamander (Ambystoma gracile) in Oregon.'' American Midland Naturalist, 109, 40-49.
Titus, T. A. and Gaines, M. S. (1991). ''Genetic variation in coastal and montane populations of Ambystoma gracile (Caudata: Ambystomatidae).'' Occasional Paper of the Museum of Natural History, University of Kansas, 141, 1-12.
Tyler, T. J., Liss, W. J., Hoffman, R. L., and Ganio, L. M. (1998). "Experimental analysis of trout effects on survival, growth, and habitat use of two species of ambystomatid salamanders." Journal of Herpetology, 32(3), 345-349.
Walls, S. C., Conrad, C. S. Murillo, M. L., and Blaustein, A. R. (1996). ''Agonistic behaviour in larvae of the Northwestern salamander (Ambystoma gracile): The effect of kinship, familiarity, and population source.'' Behaviour, 133(13-14), 965-984.
Watney, G. M. S. (1941). ''Notes on the life history of Ambystoma gracile Baird.'' Copeia, 1941, 14-17.
Weller, W. and Green, D. M. (1997). ''Checklist and current status of Canadian amphibians.'' Amphibians in Decline: Canadian Studies of a Global Problem. D. M. Green, eds., Herpetological Conservation, 309-328.
Originally submitted by: John Romansic (first posted 1999-04-13)
Edited by: Kellie Whittaker (2023-08-11)Species Account Citation: AmphibiaWeb 2023 Ambystoma gracile: Northwestern Salamander <https://amphibiaweb.org/species/3833> University of California, Berkeley, CA, USA. Accessed Apr 19, 2024.
Feedback or comments about this page.
Citation: AmphibiaWeb. 2024. <https://amphibiaweb.org> University of California, Berkeley, CA, USA. Accessed 19 Apr 2024.
AmphibiaWeb's policy on data use.
|

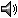

 Raffaëlli Account
Raffaëlli Account Map of Life
Map of Life